 Arg-Ala-Ala-Lys-Glu-Asn-Tyr-Ala-Leu-Ala-Ala. In the plant chloroplast, the enzyme removes the C-terminal extension of the D1 polypeptide of photosystem II
Arg-Ala-Ala-Lys-Glu-Asn-Tyr-Ala-Leu-Ala-Ala. In the plant chloroplast, the enzyme removes the C-terminal extension of the D1 polypeptide of photosystem IIContinued from EC 3.4.21.51 to EC 3.4.21.101
EC 3.4.21.101 xanthomonalisin
EC 3.4.21.102 C-terminal processing peptidase
EC 3.4.21.103 physarolisin
EC 3.4.21.104 mannan-binding lectin-associated serine protease-2
EC 3.4.21.105 rhomboid protease
EC 3.4.21.106 hepsin
EC 3.4.21.107 peptidase Do
EC 3.4.21.108 HtrA2 peptidase
EC 3.4.21.109 matriptase
EC 3.4.21.110 C5a peptidase
EC 3.4.21.111 aqualysin 1
EC 3.4.21.112 site-1 protease
EC 3.4.21.113 pestivirus NS3 polyprotein peptidase
EC 3.4.21.114 equine arterivirus serine peptidase
EC 3.4.21.115 infectious pancreatic necrosis birnavirus Vp4 peptidase
EC 3.4.21.116 SpoIVB peptidase
EC 3.4.21.117 stratum corneum chymotryptic enzyme
EC 3.4.21.118 kallikrein 8
EC 3.4.21.119 kallikrein 13
EC 3.4.21.120 oviductin
EC 3.4.21.121 Lys-Lys/Arg-Xaa endopeptidase
EC 3.4.21.122 transmembrane protease serine 2
EC 3.4.21.123 kumamolysin
EC 3.4.21.101
Accepted name: xanthomonalisin
Reaction: Cleavage of casein
Other name(s): Xanthomonas aspartic proteinase; xanthomonapepsin; sedolisin-B
Comments: Secreted by the bacterium Xanthomonas sp. Belongs In peptidase family S53.
Links to other databases: BRENDA, EXPASY, KEGG, MEROPS, Metacyc, CAS registry number: 113356-29-9
References:
1. Oda, K., Nakazima, T., Terashita, T., Suzuki, K. and Murao, S. Purification and properties of an S-PI(pepstatin Ac)-insensitive carboxyl proteinase from a Xanthomonas sp. bacterium. Agric. Biol. Chem. 51 (1987) 3073-3080.
2. Wlodawer, A., Li, M., Gustchina, A., Oyama, H., Dunn, B.M. and Oda, K. Structural and enzymatic properties of the sedolisin family of serine-carboxyl peptidases. Acta Biochim. Pol. 50 (2003) 81-102. [PMID: 12673349]
Accepted name: C-terminal processing peptidase
Reaction: The enzyme shows specific recognition of a C-terminal tripeptide, Xaa-Yaa-Zaa, in which Xaa is preferably Ala or Leu, Yaa is preferably Ala or Tyr, and Zaa is preferably Ala, but then cleaves at a variable distance from the C-terminus. A typical cleavage is -Ala-AlaArg-Ala-Ala-Lys-Glu-Asn-Tyr-Ala-Leu-Ala-Ala. In the plant chloroplast, the enzyme removes the C-terminal extension of the D1 polypeptide of photosystem II
Other names: CtpA gene product (Synechocystis sp.); photosystem II D1 protein processing peptidase; protease Re; tail-specific protease; Tsp protease
Comments: Proteolytic processing of the D1 protein of photosystem II is necessary to allow the light-driven assembly of the tetranuclear manganese cluster, which is responsible for photosynthetic water oxidation. The recognition of the substrate is mediated by a PDZ domain, a small protein module that promotes protein-protein interactions by binding to internal or C-terminal sequences of their partner proteins. Type example of peptidase family S41.
Links to other databases: BRENDA, EXPASY, KEGG, MEROPS, Metacyc, PDB, CAS registry number: 216484-75-2 and 92480-11-0
References
1. Keiler, K.C. and Sauer, R.T. Tsp protease. In: Barrett, A.J., Rawlings, N.D. and Woessner, J.F. (Eds.), Handbook of Proteolytic Enzymes, Academic Press, London, 1998, pp. 460-461.
2. Beebe, K.D., Shin, J.N., Peng, J., Chaudhury, C., Khera, J. and Pei, D.H. Substrate recognition through a PDZ domain in tail-specific protease. Biochemistry 39 (2000) 3149-3155. [PMID: 10715137]
3. Liao, D.I., Qian, J., Chisholm, D.A., Jordan, D.B. and Diner, B.A. Crystal structures of the photosystem II D1 C-terminal processing protease. Nat. Struct. Biol. 7 (2000) 749-753. [PMID: 10966643]
Accepted name: physarolisin
Reaction: Milk clotting activity. Preferential cleavage of Gly8Ser in B chain of insulin most rapidly, followed by Leu11Val, Cys(SO3H)19Gly and Phe24Phe. No action on Ac-Phe-Tyr(I)2.
Other name(s): Dictyostelium discoideum aspartic proteinase; Dictyostelium discoideum aspartic proteinase E; Physarum flavicomum aspartic proteinase; Physarum polycephalum acid proteinase; Physarum aspartic proteinase; physaropepsin
Comments: Belongs in peptidase family S53. From the slime mold Physarum polycephalum. Is not inhibited by pepstatin, but is blocked by methyl 2-diazoacetamidohexanoate. Closely similar enzymes are found in Dictyostelium discoideum and P. flavicomum. Formerly included in EC 3.4.23.6.
Links to other databases: BRENDA, EXPASY, KEGG, MEROPS, Metacyc, CAS registry number: 94949-28-7
References:
1. Henney, H.R. and Tavana, G. Purification and some properties of an intracellular acid (carboxyl) proteinase from differentiating haploid cells of Physarum flavicomum. Exp. Mycol. 6 (1982) 161-170.
2. Murakami-Murofushi, K., Hiratsuka, A. and Ohta, J. A novel acid protease from haploid amoebae of Physarum polycephalum, and its changes during mating and subsequent differentiation into diploid plasmodia. Cell Struct. Funct. 9 (1984) 311-315.
3. North, M.J. and Whyte, A. Purification and characterization of two acid proteinases from Dictyostelium discoideum. J. Gen. Microbiol. 130 (1984) 123-134.
4. Wlodawer, A., Li, M., Gustchina, A., Oyama, H., Dunn, B.M. and Oda, K. Structural and enzymatic properties of the sedolisin family of serine-carboxyl peptidases. Acta Biochim. Pol. 50 (2003) 81-102. [PMID: 12673349]
5. Nishii, W., Ueki, T., Miyashita, R., Kojima, M., Kim, Y.T., Sasaki, N., Murakami-Murofushi, K. and Takahashi, K. Structural and enzymatic characterization of physarolisin (formerly physaropepsin) proves that it is a unique serine-carboxyl proteinase. Biochem. Biophys. Res. Commun. 301 (2003) 1023-1029. [PMID: 12589815]
Accepted name: mannan-binding lectin-associated serine protease-2
Reaction: Selective cleavage after Arg223 in complement component C2 (-Ser-Leu-Gly-ArgLys-Ile-Gln-Ile) and after Arg76 in complement component C4 (-Gly-Leu-Gln-ArgAla-Leu-Glu-Ile)
Other name(s): MASP-2; MBP-associated serine protease-2; mannose-binding lectin-associated serine protease-2; p100; mannan-binding lectin-associated serine peptidase
Comments: Mannan-binding lectin (MBL) recognizes patterns of neutral carbohydrates, such as mannose and N-acetylglucosamine, on a wide range of microbial surfaces and is able to initiate activation of the lectin pathway of complement [7]. This enzyme displays C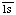 -like esterolytic activity (cf. EC 3.4.21.42, complement subcomponent C). It also cleaves C4 and C2 with efficiencies that are relatively higher than those of EC 3.4.21.42 [3]. Belongs in peptidase family S1A.
-like esterolytic activity (cf. EC 3.4.21.42, complement subcomponent C). It also cleaves C4 and C2 with efficiencies that are relatively higher than those of EC 3.4.21.42 [3]. Belongs in peptidase family S1A.
Links to other databases: BRENDA, EXPASY, KEGG, MEROPS, Metacyc, PDB, CAS registry number: 214915-16-9
References:
1. Matsushita, M. and Fujita, T. Activation of the classical complement pathway by mannose-binding protein in association with a novel C1s-like serine protease. J. Exp. Med. 176 (1992) 1497-1502. [PMID: 1460414]
2. Thiel, S., Vorup-Jensen, T., Stover, C.M., Schwaeble, W., Laursen, S.B., Poulsen, K., Willis, A.C., Eggleton, P., Hansen, S., Holmskov, U., Reid, K.B. and Jensenius, J.C. A second serine protease associated with mannan-binding lectin that activates complement. Nature 386 (1997) 506-510. [PMID: 9087411]
3. Rossi, V., Cseh, S., Bally, I., Thielens, N.M., Jensenius, J.C. and Arlaud, G.J. Substrate specificities of recombinant mannan-binding lectin-associated serine proteases-1 and -2. J Biol. Chem. 276 (2001) 40880-40887. [PMID: 11527969]
4. Ambrus, G., Gal, P., Kojima, M., Szilagyi, K., Balczer, J., Antal, J., Graf, L., Laich, A., Moffatt, B.E., Schwaeble, W., Sim, R.B. and Zavodszky, P. Natural substrates and inhibitors of mannan-binding lectin-associated serine protease-1 and -2: a study on recombinant catalytic fragments. J. Immunol. 170 (2003) 1374-1382. [PMID: 12538697]
5. Harmat, V., Gal, P., Kardos, J., Szilagyi, K., Ambrus, G., Vegh, B., Naray-Szabo, G. and Zavodszky, P. The structure of MBL-associated serine protease-2 reveals that identical substrate specificities of C1s and MASP-2 are realized through different sets of enzyme-substrate interactions. J. Mol. Biol. 342 (2004) 1533-1546. [PMID: 15364579]
6. Chen, C.B. and Wallis, R. Two mechanisms for mannose-binding protein modulation of the activity of its associated serine proteases. J. Biol. Chem. 279 (2004) 26058-26065. [PMID: 15060079]
7. Teillet, F., Dublet, B., Andrieu, J.P., Gaboriaud, C., Arlaud, G.J. and Thielens, N.M. The two major oligomeric forms of human mannan-binding lectin: chemical characterization, carbohydrate-binding properties, and interaction with MBL-associated serine proteases. J. Immunol. 174 (2005) 2870-2877. [PMID: 15728497]
Accepted name: rhomboid protease
Reaction: Cleaves type-1 transmembrane domains using a catalytic dyad composed of serine and histidine that are contributed by different transmembrane domains
Comments: These endopeptidases are multi-spanning membrane proteins. Their catalytic site is embedded within the membrane and they cleave type-1 transmembrane domains. A catalytic dyad is involved in proteolysis rather than a catalytic triad, as was thought previously [14]. They are important for embryo development in Drosophila melanogaster. Rhomboid is a key regulator of EGF receptor signalling and is responsible for cleaving Spitz, the main ligand of the Drosophila EGF receptor pathway. Belongs in peptidase family S54. Parasite-encoded rhomboid enzymes are also important for invasion of host cells by Toxoplasma and the malaria parasite. Rhomboids are widely conserved from bacteria to archaea to humans [9,13].
Links to other databases: BRENDA, EXPASY, KEGG, MEROPS, Metacyc, PDB, CAS registry number: 713145-02-9
References:
1. Urban, S. and Wolfe, M.S. Reconstitution of intramembrane proteolysis in vitro reveals that pure rhomboid is sufficient for catalysis and specificity. Proc. Natl. Acad. Sci. USA 102 (2005) 1883-1888. [PMID: 15684070]
2. Brossier, F., Jewett, T.J., Sibley, L.D. and Urban, S. A spatially localized rhomboid protease cleaves cell surface adhesins essential for invasion by Toxoplasma. Proc. Natl. Acad. Sci. USA 102 (2005) 4146-4151. [PMID: 15753289]
3. Herlan, M., Bornhovd, C., Hell, K., Neupert, W. and Reichert, A.S. Alternative topogenesis of Mgm1 and mitochondrial morphology depend on ATP and a functional import motor. J. Cell Biol. 165 (2004) 167-173. [PMID: 15096522]
4. Pascall, J.C. and Brown, K.D. Intramembrane cleavage of ephrinB3 by the human rhomboid family protease, RHBDL 2. Biochem. Biophys. Res. Commun. 317 (2004) 244–252. [PMID: 15047175]
5. Sik, A., Passer, B.J., Koonin, E.V. and Pellegrini, L. Self-regulated cleavage of the mitochondrial intramembrane-cleaving protease PARL yields Pβ, a nuclear-targeted peptide. J. Biol. Chem. 279 (2004) 15323-15329. [PMID: 14732705]
6. Urban, S. and Freeman, M. Substrate specificity of Rhomboid intramembrane proteases is governed by helix-breaking residues in the substrate transmembrane domain. Mol. Cell 11 (2003) 1425-1434. [PMID: 12820957]
7. Herlan, M., Vogel, F., Bornhovd, C., Neupert, W. and Reichert, A.S. Processing of Mgm1 by the rhomboid-type protease Pcp1 is required for maintenance of mitochondrial morphology and of mitochondrial DNA. J. Biol. Chem. 278 (2003) 27781-27788. [PMID: 12707284]
8. McQuibban, G.A., Saurya, S. and Freeman, M. Mitochondrial membrane remodelling regulated by a conserved rhomboid protease. Nature 423 (2003) 537-541. [PMID: 12774122]
9. Koonin, E.V., Makarova, K.S., Rogozin, I.B., Davidovic, L., Letellier, M.C. and Pellegrini, L. The rhomboids: a nearly ubiquitous family of intramembrane serine proteases that probably evolved by multiple ancient horizontal gene transfers. Genome Biol. 4 (2003) R19 only. [PMID: 12620104]
10. Urban, S. and Freeman, M. Intramembrane proteolysis controls diverse signalling pathways throughout evolution. Curr. Opin. Genet. Dev. 12 (2002) 512-518. [PMID: 12200155]
11. Urban, S., Schlieper, D. and Freeman, M. Conservation of intramembrane proteolytic activity and substrate specificity in prokaryotic and eukaryotic Rhomboids. Curr. Biol. 12 (2002) 1507-1512. [PMID: 12225666]
12. Urban, S., Lee, J.R. and Freeman, M. A family of Rhomboid intramembrane proteases activates all Drosophila membrane-tethered EGF-like ligands. EMBO J. 21 (2002) 4277-4286. [PMID: 12169630]
13. Urban, S., Lee, J.R. and Freeman, M. Drosophila Rhomboid-1 defines a family of putative intramembrane serine proteases. Cell 107 (2001) 173-sh;182. [PMID: 11672525]
14. Lemberg, M.K., Menendez, J., Misik, A., Garcia, M., Koth, C.M. and Freeman, M. Mechanism of intramembrane proteolysis investigated with purified rhomboid proteases. EMBO J. 24 (2005) 464-472. [PMID: 15616571]
15. Wang, Y., Zhang, Y. and Ha, Y. Crystal structure of a rhomboid family intramembrane protease. Nature 444 (2006) 179-180. [PMID: 17051161]
Accepted name: hepsin
Reaction: Cleavage after basic amino-acid residues, with Arg strongly preferred to Lys
Comments: This type-II membrane-associated serine peptidase has been implicated in cell growth and development [1,3]. The enzyme has been shown to activate blood coagulation factor VII by cleavage of the Arg152Ile153 peptide bound in BHK cells, thus indicating a possible role in the initiation of blood coagulation [2]. There is no cleavage after aromatic or aliphatic residues [1]. The occupancy of the S2 site is an absolute requirement for catalysis and a basic residue at that site is preferred to an aliphatic residue. The nature of the residue at S3 also affects hydrolysis, with Gln being much more favourable than Ala [1]. Belongs in peptidase family S1A.
Links to other databases: BRENDA, EXPASY, KEGG, MEROPS, Metacyc, PDB, CAS registry number: 112398-23-9
References:
1. Zhukov, A., Hellman, U. and Ingelman-Sundberg, M. Purification and characterization of hepsin from rat liver microsomes. Biochim. Biophys. Acta 1337 (1997) 85-95. [PMID: 9003440]
2. Kazama, Y., Hamamoto, T., Foster, D.C. and Kisiel, W. Hepsin, a putative membrane-associated serine protease, activates human factor VII and initiates a pathway of blood coagulation on the cell surface leading to thrombin formation. J. Biol. Chem. 270 (1995) 66-72. [PMID: 7814421]
3. Torres-Rosado, A., O'Shea, K.S., Tsuji, A., Chou, S.H. and Kurachi, K. Hepsin, a putative cell-surface serine protease, is required for mammalian cell growth. Proc. Natl. Acad. Sci. USA 90 (1993) 7181-7185. [PMID: 8346233]
Accepted name: peptidase Do
Reaction: Acts on substrates that are at least partially unfolded. The cleavage site P1 residue is normally between a pair of hydrophobic residues, such as ValVal
Other name(s): DegP; DegP protease; HtrA; high temperature requirement protease A; HrtA heat shock protein; protease Do; Do protease
Comments: This serine endopeptidase is essential for the clearance of denatured or aggregated proteins from the inner-membrane and periplasmic space in Escherichia coli. Natural substrates of the enzyme include colicin A lysis protein, pilin subunits and MalS from E. coli [3]. The enzyme has weak peptidase activity with casein and other non-native substrates [3]. The peptidase acts as a chaperone at low temperatures but switches to a peptidase (heat shock protein) at higher temperatures [1,6]. Molecular chaperones and peptidases control the folded state of proteins by recognizing hydrophobic stretches of polypeptide that become exposed by misfolding or unfolding. They then bind these hydrophobic substrates to prevent aggregation or assist in protein refolding. If attempts at refolding fail, then irreversibly damaged proteins are degraded by peptidases such as this enzyme [6]. Belongs in peptidase family S1C.
Links to other databases: BRENDA, EXPASY, KEGG, Metacyc, PDB, CAS registry number: 161108-11-8
References:
1. Lipinska, B., Zylicz, M. and Georgopoulos, C. The HtrA (DegP) protein, essential for Escherichia coli survival at high temperatures, is an endopeptidase. J. Bacteriol. 172 (1990) 1791-1797. [PMID: 2180903]
2. Seol, J.H., Woo, S.K., Jung, E.M., Yoo, S.J., Lee, C.S., Kim, K.J., Tanaka, K., Ichihara, A., Ha, D.B. and Chung, C.H. Protease Do is essential for survival of Escherichia coli at high temperatures: its identity with the htrA gene product. Biochem. Biophys. Res. Commun. 176 (1991) 730-736. [PMID: 2025286]
3. Jones, C.H., Dexter, P., Evans, A.K., Liu, C., Hultgren, S.J. and Hruby, D.E. Escherichia coli DegP protease cleaves between paired hydrophobic residues in a natural substrate: the PapA pilin. J. Bacteriol. 184 (2002) 5762-5771. [PMID: 12270835]
4. Swamy, K.H., Chung, C.H. and Goldberg, A.L. Isolation and characterization of protease Do from Escherichia coli, a large serine protease containing multiple subunits. Arch. Biochem. Biophys. 224 (1983) 543-554. [PMID: 6347072]
5. Pallen, M.J. and Wren, B.W. The HtrA family of serine proteases. Mol. Microbiol. 26 (1997) 209-221. [PMID: 9383148]
6. Krojer, T., Garrido-Franco, M., Huber, R., Ehrmann, M. and Clausen, T. Crystal structure of DegP (HtrA) reveals a new protease-chaperone machine. Nature 416 (2002) 455-459. [PMID: 11919638]
Accepted name: HtrA2 peptidase
Reaction: Cleavage of non-polar aliphatic amino-acids at the P1 position, with a preference for Val, Ile and Met. At the P2 and P3 positions, Arg is selected most strongly with a secondary preference for other hydrophilic residues
Other name(s): high temperature requirement protein A2; HtrA2; Omi stress-regulated endoprotease; serine proteinase OMI; HtrA2 protease; OMI/HtrA2 protease; HtrA2/Omi; Omi/HtrA2
Comments: This enzyme is upregulated in mammalian cells in response to stress induced by both heat shock and tunicamycin treatment [4]. It can induce apoptosis in a caspase-independent manner through its peptidase activity and in a caspase-dependent manner by disrupting the interaction between caspase and the inhibitor of apoptosis (IAP) [3]. Belongs in peptidase family S1C.
Links to other databases: BRENDA, EXPASY, KEGG, MEROPS, Metacyc, PDB, CAS registry number:
References:
1. Srinivasula, S.M., Gupta, S., Datta, P., Zhang, Z., Hegde, R., Cheong, N., Fernandes-Alnemri, T. and Alnemri, E.S. Inhibitor of apoptosis proteins are substrates for the mitochondrial serine protease Omi/HtrA2. J. Biol. Chem. 278 (2003) 31469-31472. [PMID: 12835328]
2. Savopoulos, J.W., Carter, P.S., Turconi, S., Pettman, G.R., Karran, E.H., Gray, C.W., Ward, R.V., Jenkins, O. and Creasy, C.L. Expression, purification, and functional analysis of the human serine protease HtrA2. Protein. Expr. Purif. 19 (2000) 227-234. [PMID: 10873535]
3. Martins, L.M., Turk, B.E., Cowling, V., Borg, A., Jarrell, E.T., Cantley, L.C. and Downward, J. Binding specificity and regulation of the serine protease and PDZ domains of HtrA2/Omi. J. Biol. Chem. 278 (2003) 49417-49427. [PMID: 14512424]
4. Gray, C.W., Ward, R.V., Karran, E., Turconi, S., Rowles, A., Viglienghi, D., Southan, C., Barton, A., Fantom, K.G., West, A., Savopoulos, J., Hassan, N.J., Clinkenbeard, H., Hanning, C., Amegadzie, B., Davis, J.B., Dingwall, C., Livi, G.P. and Creasy, C.L. Characterization of human HtrA2, a novel serine protease involved in the mammalian cellular stress response. Eur. J. Biochem. 267 (2000) 5699-5710. [PMID: 10971580]
5. Li, W., Srinivasula, S.M., Chai, J., Li, P., Wu, J.W., Zhang, Z., Alnemri, E.S. and Shi, Y. Structural insights into the pro-apoptotic function of mitochondrial serine protease HtrA2/Omi. Nat. Struct. Biol. 9 (2002) 436-441. [PMID: 11967569]
Accepted name: matriptase
Reaction: Cleaves various synthetic substrates with Arg or Lys at the P1 position and prefers small side-chain amino acids, such as Ala and Gly, at the P2 position
Other name(s): serine protease 14; membrane-type serine protease 1; MT-SP1; prostamin; serine protease TADG-15; tumor-associated differentially-expressed gene 15 protein; ST14; breast cancer 80 kDa protease; epithin; serine endopeptidase SNC19; matriptase-1; matriptase-2; matriptase-3; TMPRSS6 (gene name)
Comments: This trypsin-like integral-membrane serine peptidase has been implicated in breast cancer invasion and metastasis [1,2]. The enzyme can activate hepatocyte growth factor/scattering factor (HGF/SF) by cleavage of the two-chain forms at an Arg residue to give active α- and β-HGF, but It does not activate plasminogen, which shares high homology with HGF [1]. The enzyme can also activate urokinase plasminogen activator (uPA), which initiates the matrix-degrading peptidase cascade [1,2]. Hemojuvelin has been shown to be a physiological substrate for matriptase-2 [5]. Belongs in peptidase family S1A.
Links to other databases: BRENDA, EXPASY, KEGG, MEROPS, MetaCyc, PDB, CAS registry number: 241475-96-7
References:
1. Lee, S.L., Dickson, R.B. and Lin, C.Y. Activation of hepatocyte growth factor and urokinase/plasminogen activator by matriptase, an epithelial membrane serine protease. J. Biol. Chem. 275 (2000) 36720-36725. [PMID: 10962009]
2. Lin, C.Y., Anders, J., Johnson, M., Sang, Q.A. and Dickson, R.B. Molecular cloning of cDNA for matriptase, a matrix-degrading serine protease with trypsin-like activity. J. Biol. Chem. 274 (1999) 18231-18236. [PMID: 10373424]
3. Ramsay, A.J., Reid, J.C., Velasco, G., Quigley, J.P. and Hooper, J.D. The type II transmembrane serine protease matriptase-2 – identification, structural features, enzymology, expression pattern and potential roles. Front. Biosci. 13 (2008) 569-579. [PMID: 17981570]
4. Kojima, K., Tsuzuki, S., Fushiki, T. and Inouye, K. The activity of a type II transmembrane serine protease, matriptase, is dependent solely on the catalytic domain. Biosci. Biotechnol. Biochem. 73 (2009) 454-456. [PMID: 19202271]
5. Wysocka, M., Gruba, N., Miecznikowska, A., Popow-Stellmaszyk, J., Gutschow, M., Stirnberg, M., Furtmann, N., Bajorath, J., Lesner, A. and Rolka, K. Substrate specificity of human matriptase-2. Biochimie 97 (2014) 121-127. [PMID: 24161741]
Accepted name: C5a peptidase
Reaction: The primary cleavage site is at His67Lys68 in human C5a with a minor secondary cleavage site at Ala58Ser59
Other name(s): streptococcal C5a peptidase; ScpA; ScpB; SCPA
Comments: This enzyme is a surface-associated subtilisin-like serine peptidase with very specific substrate specificity. Virulent strains of streptococci, including Streptococcus pyogenes, can evade human detection and phagocytosis by destroying the complement chemotaxin C5a. Cleavage of human C5a by this enzyme reduces the ability of C5a to bind receptors on the surface of polymorphonuclear neutrophil leukocytes (PMNLs) and thereby abolishes its chemotactic properties [1,4]. Belongs in peptidase family S8A.
Links to other databases: BRENDA, EXPASY, KEGG, MEROPS, Metacyc, PDB, CAS registry number: 100179-39-3
References:
1. Wexler, D.E., Chenoweth, D.E. and Cleary, P.P. Mechanism of action of the group A streptococcal C5a inactivator. Proc. Natl. Acad. Sci. USA 82 (1985) 8144-8148. [PMID: 3906656]
2. Bohnsack, J.F., Mollison, K.W., Buko, A.M., Ashworth, J.C. and Hill, H.R. Group B streptococci inactivate complement component C5a by enzymic cleavage at the C-terminus. Biochem. J. 273 (1991) 635-640. [PMID: 1996961]
3. Cleary, P.P., Prahbu, U., Dale, J.B., Wexler, D.E. and Handley, J. Streptococcal C5a peptidase is a highly specific endopeptidase. Infect. Immun. 60 (1992) 5219-5223. [PMID: 1452354]
4. Anderson, E.T., Wetherell, M.G., Winter, L.A., Olmsted, S.B., Cleary, P.P. and Matsuka, Y.V. Processing, stability, and kinetic parameters of C5a peptidase from Streptococcus pyogenes. Eur. J. Biochem. 269 (2002) 4839-4851. [PMID: 12354115]
5. Stafslien, D.K. and Cleary, P.P. Characterization of the streptococcal C5a peptidase using a C5a-green fluorescent protein fusion protein substrate. J. Bacteriol. 182 (2000) 3254-3258. [PMID: 10809707]
6. Terao, Y., Yamaguchi, M., Hamada, S. and Kawabata, S. Multifunctional glyceraldehyde-3-phosphate dehydrogenase of Streptococcus pyogenes is essential for evasion from neutrophils. J. Biol. Chem. 281 (2006) 14215-14223. [PMID: 16565520]
Accepted name: aqualysin 1
Reaction: Exhibits low specificity towards esters of amino acids with small hydrophobic or aromatic residues at the P1 position
Other name(s): caldolysin
Comments: This enzyme from the extreme thermophile, Thermus aquaticus, is an alkaline serine peptidase. It has three subsites, S1, S2, and S3, in the substrate binding site. The preferred amino acids at the S1 site are Ala and Phe, at the S2 site are Ala and norleucine and at the S3 site are Phe and Ile [3]. These specificities are similar to those of EC 3.4.21.64 (peptidase K) and EC 3.4.21.62 (subtilisin BPN') [3]. The enzyme displays broad specificity for cleavage of insulin B-chain and hydrolyses elastin substrates such as succinyl-(Ala)n-p-nitroanilide (n = 1,2,3) and some peptide esters [1,3]. Belongs in peptidase family S8A.
Links to other databases: BRENDA, EXPASY, KEGG, MEROPS, Metacyc, PDB, CAS registry number: 88747-68-6
References:
1. Matsuzawa, H., Tokugawa, K., Hamaoki, M., Mizoguchi, M., Taguchi, H., Terada, I., Kwon, S.T. and Ohta, T. Purification and characterization of aqualysin I (a thermophilic alkaline serine protease) produced by Thermus aquaticus YT-1. Eur. J. Biochem. 171 (1988) 441-447. [PMID: 3162211]
2. Tanaka, T., Matsuzawa, H., Kojima, S., Kumagai, I., Miura, K. and Ohta, T. P1 specificity of aqualysin I (a subtilisin-type serine protease) from Thermus aquaticus YT-1, using P1-substituted derivatives of Streptomyces subtilisin inhibitor. Biosci. Biotechnol. Biochem. 62 (1998) 2035-2038. [PMID: 9882104]
3. Tanaka, T., Matsuzawa, H. and Ohta, T. Substrate specificity of aqualysin I, a bacterial thermophilic alkaline serine protease from Thermus aquaticus YT-1: Comparison with proteinase K, subtilisin BPN' and subtilisin Carlsberg. Biosci. Biotechnol. Biochem. 62 (1998) 2161-2165.
Accepted name: site-1 protease
Reaction: Processes precursors containing basic and hydrophobic/aliphatic residues at P4 and P2, respectively, with a relatively relaxed acceptance of amino acids at P1 and P3
Other name(s): mammalian subtilisin/kexin isozyme 1; membrane-bound transcription factor site-1 protease; proprotein convertase SKI-1; proprotein convertase SKI-1/S1PPS1; S1P endopeptidase; S1P protease; site-1 peptidase; site-1 protease; SKI-1; SREBP proteinase; SREBP S1 protease; SREBP-1 proteinase; SREBP-2 proteinase; sterol regulatory element-binding protein proteinase; sterol regulatory element-binding protein site 1 protease; sterol-regulated luminal protease; subtilase SKI-1; subtilase SKI-1/S1P; subtilisin/kexin-isozyme 1
Comments: Cleaves sterol regulatory element-binding proteins (SREBPs) and thereby initiates a process by which the active fragments of the SREBPs translocate to the nucleus and activate genes controlling the synthesis and uptake of cholesterol and unsaturated fatty acids into the bloodstream [1]. The enzyme also processes pro-brain-derived neurotrophic factor and undergoes autocatalytic activation in the endoplasmic reticulum through sequential cleavages [5]. The enzyme can also process the unfolded protein response stress factor ATF6 at an Arg-His-Lys-Lys site [4,8], and the envelope glycoprotein of the highly infectious Lassa virus [5,8] and Crimean Congo hemorrhagic fever virus at Arg-Arg-Lys-Lys [7,8]. Belongs in peptidase family S8A.
Links to other databases: BRENDA, EXPASY, KEGG, MEROPS, Metacyc, CAS registry number: 167140-48-9
References:
1. Espenshade, P.J., Cheng, D., Goldstein, J.L. and Brown, M.S. Autocatalytic processing of site-1 protease removes propeptide and permits cleavage of sterol regulatory element-binding proteins. J. Biol. Chem. 274 (1999) 22795-22804. [PMID: 10428864]
2. Cheng, D., Espenshade, P.J., Slaughter, C.A., Jaen, J.C., Brown, M.S. and Goldstein, J.L. Secreted site-1 protease cleaves peptides corresponding to luminal loop of sterol regulatory element-binding proteins. J. Biol. Chem. 274 (1999) 22805-22812. [PMID: 10428865]
3. Touré, B.B., Munzer, J.S., Basak, A., Benjannet, S., Rochemont, J., Lazure, C., Chrétien, M. and Seidah, N.G. Biosynthesis and enzymatic characterization of human SKI-1/S1P and the processing of its inhibitory prosegment. J. Biol. Chem. 275 (2000) 2349-2358. [PMID: 10644685]
4. Ye, J., Rawson, R.B., Komuro, R., Chen, X., Dave, U.P., Prywes, R., Brown, M.S. and Goldstein, J.L. ER stress induces cleavage of membrane-bound ATF6 by the same proteases that process SREBPs. Mol. Cell 6 (2000) 1355-1364. [PMID: 11163209]
5. Lenz, O., ter Meulen, J., Klenk, H.D., Seidah, N.G. and Garten, W. The Lassa virus glycoprotein precursor GP-C is proteolytically processed by subtilase SKI-1/S1P. Proc. Natl. Acad. Sci. USA 98 (2001) 12701-12705. [PMID: 11606739]
6. Basak, A., Chrétien, M. and Seidah, N.G. A rapid fluorometric assay for the proteolytic activity of SKI-1/S1P based on the surface glycoprotein of the hemorrhagic fever Lassa virus. FEBS Lett. 514 (2002) 333-339. [PMID: 11943176]
7. Vincent, M.J., Sanchez, A.J., Erickson, B.R., Basak, A., Chretien, M., Seidah, N.G. and Nichol, S.T. Crimean-Congo hemorrhagic fever virus glycoprotein proteolytic processing by subtilase SKI-1. J. Virol. 77 (2003) 8640-8649. [PMID: 12885882]
8. Seidah, N.G. and Chrétien, M. Proprotein convertase SKI-1/S1P. In: Barrett, A.J., Rawlings, N.D. and Woessner, J.F. (Eds), Handbook of Proteolytic Enzymes, 2nd edn, vol. 2, Elsevier, London, 2004, pp. 1845-1847.
Accepted name: pestivirus NS3 polyprotein peptidase
Reaction: Leu is conserved at position P1 for all four cleavage sites. Alanine is found at position P1' of the NS4A-NS4B cleavage site, whereas serine is found at position P1' of the NS3-NS4A, NS4B-NS5A and NS5A-NS5B cleavage sites
Other name(s): border disease virus NS3 endopeptidase; BDV NS3 endopeptidase; bovine viral diarrhea virus NS3 endopeptidase; BVDV NS3 endopeptidase; classical swine fever virus NS3 endopeptidase; CSFV NS3 endopeptidase; p80
Comments: The polyprotein of noncytopathogenic pestiviruses is cleaved co- and post-translationally into at least 11 proteins (Npro, C, Erns, E1, E2, p7, NS2-3, NS4A, NS4B, NS5A, and NS5B) [2]. The genomes of cytopathogenic pestivirus strains express at least one additional protein, called NS3 (p80) [2]. This enzyme, which resides in the N-terminal region of NS3 (nonstructural protein 3), is essential for generation of its own C-terminus and for processing of the downstream cleavage sites, leading to the release of the pestivirus nonstructural proteins NS4A, NS4B, NS5A and NS5B [1,2]. Belongs in peptidase family S31.
Links to other databases: BRENDA, EXPASY, KEGG, MEROPS, Metacyc, PDB, CAS registry number:
References:
1. Wiskerchen, M. and Collett, M.S. Pestivirus gene expression: protein p80 of bovine viral diarrhea virus is a proteinase involved in polyprotein processing. Virology 184 (1991) 341-350. [PMID: 1651596]
2. Tautz, N., Elbers, K., Stoll, D., Meyers, G. and Thiel, H.J. Serine protease of pestiviruses: determination of cleavage sites. J. Virol. 71 (1997) 5415-5422. [PMID: 9188613]
3. Xu, J., Mendez, E., Caron, P.R., Lin, C., Murcko, M.A., Collett, M.S. and Rice, C.M. Bovine viral diarrhea virus NS3 serine proteinase: polyprotein cleavage sites, cofactor requirements, and molecular model of an enzyme essential for pestivirus replication. J. Virol. 71 (1997) 5312-5322. [PMID: 9188600]
4. Tautz, N., Kaiser, A. and Thiel, H.J. NS3 serine protease of bovine viral diarrhea virus: characterization of active site residues, NS4A cofactor domain, and protease-cofactor interactions. Virology 273 (2000) 351-363. [PMID: 10915606]
Accepted name: equine arterivirus serine peptidase
Reaction: Cleavage of (Glu/Gln)(Gly/Ser/Ala) in arterivirus replicase translation products ORF1a and ORF1ab
Glossary: arterivirus nsp4; equine arteritis virus serine peptidase; 3C-like serine protease; 3C-like Ser protease; 3CLSP; nonstructural protein 4 serine protease, Nsp4 serine protease; nsp4 serine protease; Nsp4 SP; chymotrypsin-like serine proteinase nsp4
Comments: In the equine arterivirus (EAV), the replicase gene is translated into open reading frame 1a (ORF1a) and ORF1ab polyproteins. This enzyme is the main viral proteinase and processes five cleavage sites in the ORF1a protein and three in the ORF1b-encoded part of the ORF1ab protein to yield nonstructural proteins (nsp5-nsp12) [3]. It combines the catalytic system of a chymotrypsin-like serine peptidase (His-Asp-Ser catalytic triad) with the substrate specificity of a 3C-like serine peptidase (Glu or Gln) at the P1 position and a small amino-acid residue (Gly, Ser or Ala) at the P1' position [1]. Cleavage of ORF1ab by this enzyme is essential for viral replication [2]. Belongs in peptidase family S32.
Links to other databases: BRENDA, EXPASY, KEGG, MEROPS, Metacyc, PDB, CAS registry number:
References:
1. Snijder, E.J., Wassenaar, A.L., van Dinten, L.C., Spaan, W.J. and Gorbalenya, A.E. The arterivirus nsp4 protease is the prototype of a novel group of chymotrypsin-like enzymes, the 3C-like serine proteases. J. Biol. Chem. 271 (1996) 4864-4871. [PMID: 8617757]
2. van Dinten, L.C., Rensen, S., Gorbalenya, A.E. and Snijder, E.J. Proteolytic processing of the open reading frame 1b-encoded part of arterivirus replicase is mediated by nsp4 serine protease and is essential for virus replication. J. Virol. 73 (1999) 2027-2037. [PMID: 9971783]
3. Barrette-Ng, I.H., Ng, K.K., Mark, B.L., Van Aken, D., Cherney, M.M., Garen, C., Kolodenko, Y., Gorbalenya, A.E., Snijder, E.J. and James, M.N. Structure of arterivirus nsp4. The smallest chymotrypsin-like proteinase with an α/β C-terminal extension and alternate conformations of the oxyanion hole. J. Biol. Chem. 277 (2002) 39960-39966. [PMID: 12163505]
Accepted name: infectious pancreatic necrosis birnavirus Vp4 peptidase
Reaction: Cleaves the (Ser/Thr)-Xaa-Ala(Ser/Ala)-Gly motif in the polyprotein NH2-pVP2-VP4-VP3-COOH of infectious pancreatic necrosis virus at the pVP2-VP4 and VP4-VP3 junctions
Other name(s): infectious pancreatic necrosis virus protease; IPNV Vp4 protease; IPNV Vp4 peptidase; NS protease; NS-associated protease; Vp4 protease
Comments: Infectious pancreatic necrosis virus (IPNV) is a birnavirus that causes an acute, contagious disease in young salmonid fish [2]. As with most viruses that infect eukaryotic cells, the proteolytic processing of viral precursor proteins is a crucial step in the life cycle of this virus [2]. pVP2 is converted into VP2 by cleavage near the carboxy end of pVP2. This cleavage is most likely due to host-cell proteases rather than VP4 [2,3]. Differs from most serine peptidases in not having the catalytic triad Ser-His-Asp [2]. Belongs in peptidase family S50.
Links to other databases: BRENDA, EXPASY, KEGG, MEROPS, Metacyc, PDB, CAS registry number:
References:
1. Manning, D.S. and Leong, J.C. Expression in Escherichia coli of the large genomic segment of infectious pancreatic necrosis virus. Virology 179 (1990) 16-25. [PMID: 2219718]
2. Petit, S., Lejal, N., Huet, J.C. and Delmas, B. Active residues and viral substrate cleavage sites of the protease of the birnavirus infectious pancreatic necrosis virus. J. Virol. 74 (2000) 2057-2066. [PMID: 10666235]
3. Dobos, P. The molecular biology of infectious pancreatic necrosis virus (IPNV). Annu. Rev. Fish Dis. 5 (1995) 25-54.
Accepted name: SpoIVB peptidase
Reaction: Self-cleaves Val52Asn53, Ala62Phe63 and Val74Thr75 at the N-terminus of SpoIVB
Other name(s): sporulation factor IV B protease
Comments: This enzyme plays a central role in a regulatory checkpoint (the σK checkpoint), which coordinates gene expression during the later stages of spore formation in Bacillus subtilis [1,3]. The enzyme activates proteolytic processing of a sporulation-specific sigma factor, pro-σK, to its mature and active form, σK, by self-cleavage [1,3]. The enzyme is also subject to secondary proteolysis, which presumably inactivates SpoIVB [3]. The enzyme is also essential for the formation of heat-resistant spores. Belongs in peptidase family S55.
Links to other databases: BRENDA, EXPASY, KEGG, MEROPS, Metacyc, CAS registry number: 296241-18-4
References:
1. Wakeley, P.R., Dorazi, R., Hoa, N.T., Bowyer, J.R. and Cutting, S.M. Proteolysis of SpolVB is a critical determinant in signalling of pro-σK processing in Bacillus subtilis. Mol. Microbiol. 36 (2000) 1336-1348. [PMID: 10931284]
2. Hoa, N.T., Brannigan, J.A. and Cutting, S.M. The PDZ domain of the SpoIVB serine peptidase facilitates multiple functions. J. Bacteriol. 183 (2001) 4364-4373. [PMID: 11418578]
3. Hoa, N.T., Brannigan, J.A. and Cutting, S.M. The Bacillus subtilis signaling protein SpoIVB defines a new family of serine peptidases. J. Bacteriol. 184 (2002) 191-199. [PMID: 11741860]
4. Dong, T.C. and Cutting, S.M. SpoIVB-mediated cleavage of SpoIVFA could provide the intercellular signal to activate processing of pro-σK in Bacillus subtilis. Mol. Microbiol. 49 (2003) 1425-1434. [PMID: 12940997]
Accepted name: stratum corneum chymotryptic enzyme
Reaction: Cleavage of proteins with aromatic side chains in the P1 position
Other name(s): kallikrein 7; SCCE; KLK7; PRSS6; hK7
Comments: This enzyme has wide substrate specificity, being able to degrade heat-denatured bovine casein and the α-chain of native human fibrinogen. It cleaves the B chain of bovine insulin at Leu6Cya7, Tyr16Leu17, Phe25Tyr26 and Tyr26Thr27 [1]. It is thought to play a role in the desquamation (skin-shedding) of the outer layer of skin, the stratum corneum, by degrading intercellular cohesive structures [1,2]. Belongs in peptidase family S1A.
Links to other databases: BRENDA, EXPASY, KEGG, MEROPS, Metacyc, PDB, CAS registry number: 155215-90-0
References:
1. Skytt, A., Strömqvist, M. and Egelrud, T. Primary substrate specificity of recombinant human stratum corneum chymotryptic enzyme. Biochem. Biophys. Res. Commun. 211 (1995) 586-589. [PMID: 7794273]
2. Egelrud, T. Purification and preliminary characterization of stratum corneum chymotryptic enzyme: a proteinase that may be involved in desquamation. J. Invest. Dermatol. 101 (1993) 200-204. [PMID: 8393902]
3. Hansson, L., Strömqvist, M., Bäckman, A., Wallbrandt, P., Carlstein, A. and Egelrud, T. Cloning, expression, and characterization of stratum corneum chymotryptic enzyme. A skin-specific human serine proteinase. J. Biol. Chem. 269 (1994) 19420-19426. [PMID: 8034709]
4. Yousef, G.M., Scorilas, A., Magklara, A., Soosaipillai, A. and Diamandis, E.P. The KLK7 (PRSS6) gene, encoding for the stratum corneum chymotryptic enzyme is a new member of the human kallikrein gene family - genomic characterization, mapping, tissue expression and hormonal regulation. Gene 254 (2000) 119-128. [PMID: 10974542]
5. Vasilopoulos, Y., Cork, M.J., Murphy, R., Williams, H.C., Robinson, D.A., Duff, G.W., Ward, S.J. and Tazi-Ahnini, R. Genetic association between an AACC insertion in the 3'UTR of the stratum corneum chymotryptic enzyme gene and atopic dermatitis. J. Invest. Dermatol. 123 (2004) 62-66. [PMID: 15191543]
Accepted name: kallikrein 8
Reaction: Cleavage of amide substrates following the basic amino acids Arg or Lys at the P1 position, with a preference for Arg over Lys
Other name(s): KLK8; PRSS19; human kallikrein 8; hK8; mK8; ovasin; tumor-associated differentially expressed gene 14; TADG-14; NP; neuropsin
Comments: The enzyme is activated by removal of an N-terminal prepropeptide [2,4]. The highest amidolytic activity is observed using Boc-Val-Pro-Arg7-amido-4-methylcoumarin, which is a substrate of α-thrombin [2,4]. Substrates lacking basic amino acids in the P1 position are not cleaved [4]. The enzyme degrades casein, fibronectin, gelatin, collagen type IV, fibrinogen, and high-molecular-mass kininogen [3] and is associated with diseases such as ovarian cancer and Alzheimer's disease [4]. Belongs in peptidase family S1A.
Links to other databases: BRENDA, EXPASY, KEGG, MEROPS, Metacyc, PDB, CAS registry number: 171715-15-4
References:
1. Chen, Z.L., Yoshida, S., Kato, K., Momota, Y., Suzuki, J., Tanaka, T., Ito, J., Nishino, H., Aimoto, S., Kiyama, H. and Shiosaka, S. Expression and activity-dependent changes of a novel limbic-serine protease gene in the hippocampus. J. Neurosci. 15 (1995) 5088-5097. [PMID: 7623137]
2. Shimizu, C., Yoshida, S., Shibata, M., Kato, K., Momota, Y., Matsumoto, K., Shiosaka, T., Midorikawa, R., Kamachi, T., Kawabe, A. and Shiosaka, S. Characterization of recombinant and brain neuropsin, a plasticity-related serine protease. J. Biol. Chem. 273 (1998) 11189-11196. [PMID: 9556608]
3. Rajapakse, S., Ogiwara, K., Takano, N., Moriyama, A. and Takahashi, T. Biochemical characterization of human kallikrein 8 and its possible involvement in the degradation of extracellular matrix proteins. FEBS Lett. 579 (2005) 6879-6884. [PMID: 16337200]
4. Kishi, T., Cloutier, S.M., Kündig, C., Deperthes, D. and Diamandis, E.P. Activation and enzymatic characterization of recombinant human kallikrein 8. Biol. Chem. 387 (2006) 723-731. [PMID: 16800733]
Accepted name: kallikrein 13
Reaction: Hydrolyses mouse Ren2 protein (a species of prorenin present in the submandibular gland) on the carboxy side of the arginine residue at the Lys-Arg pair in the N-terminus, to yield mature renin
Other name(s): KLK13; kallikrein mK13; mGK-13; mK13; mKLK13; prorenin converting enzyme 1; PRECE-1; prorenin-converting enzyme; PRECE; proteinase P
Comments: The enzyme is specific for prorenin from the mouse submandibular gland, as prorenin from the mouse kidney (Ren1) and human prorenin are not substrates [1]. Site-directed mutagenesis studies have shown that the enzyme will also cleave prorenin when Lys-Arg is replaced by Arg-Arg or Gln-Arg but the rate of reaction is much slower when Lys-Lys is used. This enzyme is also able to process pro-interleukin-1β (pro-IL-1β) in mouse submandibular gland to form IL-1β [4]. Belongs in peptidase family S1A.
Links to other databases: BRENDA, EXPASY, KEGG, MEROPS, Metacyc, PDB, CAS registry number: 342900-44-1
References:
1. Nakayama, K., Kim, W.S., Nakagawa, T., Nagahama, M. and Murakami, K. Substrate specificity of prorenin converting enzyme of mouse submandibular gland. Analysis using site-directed mutagenesis. J. Biol. Chem. 265 (1990) 21027-21031. [PMID: 2250008]
2. Kim, W.S., Hatsuzawa, K., Ishizuka, Y., Hashiba, K., Murakami, K. and Nakayama, K. A processing enzyme for prorenin in mouse submandibular gland. Purification and characterization. J. Biol. Chem. 265 (1990) 5930-5933. [PMID: 2180937]
3. Kikkawa, Y., Yamanaka, N., Tada, J., Kanamori, N., Tsumura, K. and Hosoi, K. Prorenin processing and restricted endoproteolysis by mouse tissue kallikrein family enzymes (mK1, mK9, mK13, and mK22). Biochim. Biophys. Acta 1382 (1998) 55-64. [PMID: 9507064]
4. Yao, C., Karabasil, M.R., Purwanti, N., Li, X., Akamatsu, T., Kanamori, N. and Hosoi, K. Tissue kallikrein mK13 is a candidate processing enzyme for the precursor of interleukin-1β in the submandibular gland of mice. J. Biol. Chem. 281 (2006) 7968-7976. [PMID: 16423834]
Accepted name: oviductin
Reaction: Preferential cleavage at Gly-Ser-Arg373 of glycoprotein gp43 in Xenopus laevis coelemic egg envelope to yield gp41
Other name(s): oviductal protease
Comments: The egg envelope of the South African clawed frog (Xenopus laevis) is modified during transit of the egg through the pars rectus oviduct, changing the egg envelope from an unfertilizable form to a fertilizable form. This process involves the conversion of glycoprotein p43 to p41 by the pars recta protease oviductin. It is thought that the enzymatically active protease molecule comprises the N-terminal protease domain coupled to two C-terminal CUB domains, which are related to the mammalian spermadhesin molecules implicated in mediating sperm-envelope interactions [2]. The enzyme is also found in the Japanese toad (Bufo japonicus) [3]. Belongs in peptidase family S1.
Links to other databases: BRENDA, EXPASY, KEGG, MEROPS, Metacyc, CAS registry number: 342900-44-1
References:
1. Hardy, D.M. and Hedrick, J.L. Oviductin. Purification and properties of the oviductal protease that processes the molecular weight 43,000 glycoprotein of the Xenopus laevis egg envelope. Biochemistry 31 (1992) 4466-4472. [PMID: 1581303]
2. Lindsay, L.L., Wieduwilt, M.J. and Hedrick, J.L. Oviductin, the Xenopus laevis oviductal protease that processes egg envelope glycoprotein gp43, increases sperm binding to envelopes, and is translated as part of an unusual mosaic protein composed of two protease and several CUB domains. Biol. Reprod. 60 (1999) 989-995. [PMID: 10084976]
3. Hiyoshi, M., Takamune, K., Mita, K., Kubo, H., Sugimoto, Y. and Katagiri, C. Oviductin, the oviductal protease that mediates gamete interaction by affecting the vitelline coat in Bufo japonicus: its molecular cloning and analyses of expression and posttranslational activation. Dev. Biol. 243 (2002) 176-184. [PMID: 11846486]
Accepted name: Lys-Lys/Arg-Xaa endopeptidase
Reaction: Cleavage of -Lys-Lys┼ and -Lys-Arg┼ bonds.
Other name(s): ASP (Aeromonas sobria)-type peptidase; Aeromonas extracellular serine protease
Comments: The enzyme is a serine peptidase, which has been shown to cleave prothrombin and prekallikrein. It hydrolyses the complement component C5 releasing complement component C5a.
Links to other databases: BRENDA, EXPASY, KEGG, Metacyc, CAS registry number:
References:
1. Kobayashi, H., Utsunomiya, H., Yamanaka, H., Sei, Y., Katunuma, N., Okamoto, K. and Tsuge, H. Structural basis for the kexin-like serine protease from Aeromonas sobria as sepsis-causing factor. J. Biol. Chem. 284 (2009) 27655-27663. [PMID: 19654332]
2. Nitta, H., Kobayashi, H., Irie, A., Baba, H., Okamoto, K. and Imamura, T. Activation of prothrombin by ASP, a serine protease released from Aeromonas sobria. FEBS Lett 581 (2007) 5935-5939. [PMID: 18067862]
3. Kobayashi, H., Takahashi, E., Oguma, K., Fujii, Y., Yamanaka, H., Negishi, T., Arimoto-Kobayashi, S., Tsuji, T. and Okamoto, K. Cleavage specificity of the serine protease of Aeromonas sobria, a member of the kexin family of subtilases. FEMS Microbiol. Lett. 256 (2006) 165-170. [PMID: 16487335]
4. Imamura, T., Nitta, H., Wada, Y., Kobayashi, H. and Okamoto, K. Impaired plasma clottability induction through fibrinogen degradation by ASP, a serine protease released from Aeromonas sobria. FEMS Microbiol. Lett. 284 (2008) 35-42. [PMID: 18462393]
5. Nitta, H., Imamura, T., Wada, Y., Irie, A., Kobayashi, H., Okamoto, K. and Baba, H. Production of C5a by ASP, a serine protease released from Aeromonas sobria. J. Immunol. 181 (2008) 3602-3608. [PMID: 18714034]
Accepted name: transmembrane protease serine 2
Reaction: The enzyme cleaves angiotensin-converting enzyme 2 (EC 3.4.17.23) and cleaves influenzea A and B virus and coronavirus spike glycoproteins at arginine residues.
Other name(s): TMPRSS2 (gene name); epitheliasin
Comments: The enzyme, present in mammalia, cleaves and inactivates EC 3.4.17.23, angiotensin-converting enzyme 2 (ACE2) at arginine residues in the region R697 to R716, which enhances influenza and coronavirus uptake [7] The enzyme also cleaves and activates influenza and coronavirus spike glycoproteins and thus facilitates virus-cell membrane fusions. The cleavage of SARS-COV2 spike glycoprotein occurs between the S2 and S2' site at SKPSKR/SFIEDL, while the cleavage of MERS-COV glycoprotein occurs at GSRSAR/SAIEDL
Links to other databases: BRENDA, EXPASY, KEGG, MEROPS, MetaCyc, PDB, CAS registry number:
References:
1. Jacquinet, E., Rao, N.V., Rao, G.V. and Hoidal, J.R. Cloning, genomic organization, chromosomal assignment and expression of a novel mosaic serine proteinase: epitheliasin. FEBS Lett. 468 (2000) 93-100. [PMID: 10683448]
2. Jacquinet, E., Rao, N.V., Rao, G.V., Zhengming, W., Albertine, K.H. and Hoidal, J.R. Cloning and characterization of the cDNA and gene for human epitheliasin. Eur. J. Biochem. 268 (2001) 2687-2699. [PMID: 11322890]
3. Bottcher-Friebertshauser, E., Freuer, C., Sielaff, F., Schmidt, S., Eickmann, M., Uhlendorff, J., Steinmetzer, T., Klenk, H.D. and Garten, W. Cleavage of influenza virus hemagglutinin by airway proteases TMPRSS2 and HAT differs in subcellular localization and susceptibility to protease inhibitors. J. Virol. 84 (2010) 5605-5614. [PMID: 20237084]
4. Bertram, S., Glowacka, I., Muller, M.A., Lavender, H., Gnirss, K., Nehlmeier, I., Niemeyer, D., He, Y., Simmons, G., Drosten, C., Soilleux, E.J., Jahn, O., Steffen, I. and Pohlmann, S. Cleavage and activation of the severe acute respiratory syndrome coronavirus spike protein by human airway trypsin-like protease. J. Virol. 85 (2011) 13363-13372. [PMID: 21994442]
5. Bertram, S., Dijkman, R., Habjan, M., Heurich, A., Gierer, S., Glowacka, I., Welsch, K., Winkler, M., Schneider, H., Hofmann-Winkler, H., Thiel, V. and Pohlmann, S. TMPRSS2 activates the human coronavirus 229E for cathepsin-independent host cell entry and is expressed in viral target cells in the respiratory epithelium. J. Virol. 87 (2013) 6150-6160. [PMID: 23536651]
6. Abe, M., Tahara, M., Sakai, K., Yamaguchi, H., Kanou, K., Shirato, K., Kawase, M., Noda, M., Kimura, H., Matsuyama, S., Fukuhara, H., Mizuta, K., Maenaka, K., Ami, Y., Esumi, M., Kato, A. and Takeda, M. TMPRSS2 is an activating protease for respiratory parainfluenza viruses. J. Virol. 87 (2013) 11930-11935. [PMID: 23966399]
7. Heurich, A., Hofmann-Winkler, H., Gierer, S., Liepold, T., Jahn, O. and Pohlmann, S. TMPRSS2 and ADAM17 cleave ACE2 differentially and only proteolysis by TMPRSS2 augments entry driven by the severe acute respiratory syndrome coronavirus spike protein. J. Virol. 88 (2014) 1293-1307. [PMID: 24227843]
8. Limburg, H., Harbig, A., Bestle, D., Stein, D.A., Moulton, H.M., Jaeger, J., Janga, H., Hardes, K., Koepke, J., Schulte, L., Koczulla, A.R., Schmeck, B., Klenk, H.D. and Bottcher-Friebertshauser, E. TMPRSS2 is the major activating protease of influenza A virus in primary human airway cells and influenza B virus in human type II pneumocytes. J. Virol. 93 (2019) . [PMID: 31391268]
9. Bestle, D., Heindl, M.R., Limburg, H., Van Lam van, T., Pilgram, O., Moulton, H., Stein, D.A., Hardes, K., Eickmann, M., Dolnik, O., Rohde, C., Klenk, H.D., Garten, W., Steinmetzer, T. and Bottcher-Friebertshauser, E. TMPRSS2 and furin are both essential for proteolytic activation of SARS-CoV-2 in human airway cells. Life Sci Alliance 3 (2020) . [PMID: 32703818]
Accepted name: kumamolysin
Reaction: The enzyme preferentially hydrolyses peptides having an Ala or Pro residue at P2 position and prefers such charged amino acid residues as Glu or Arg at the P2' position. In the oxidized insulin B chain, kumamolysin preferentially cleaves between Leu15 and Tyr16
Other name(s): KSCP; kumamolisin
Comments: This bacterial pepstatin-insensitive carboxyl proteinase has been isolated and characterized from Bacillus sp. MN-32 and from several Burkholderia spp. Kumamolysin from Bacillus sp. MN-32 exhibits a Ser278/Glu78/Asp82 catalytic triad. The enzyme is a type example of peptidase family S53 in the MEROPS Peptidas Database.
Links to other databases: BRENDA, EXPASY, KEGG, MEROPS, MetaCyc, CAS registry number:
References:
1. Murao, S., Ohkuni, K., Nagao, M., Hirayama, K., Fukuhara, K., Oda, K., Oyama, H. and Shin, T. Purification and characterization of kumamolysin, a novel thermostable pepstatin-insensitive carboxyl proteinase from Bacillus novosp. MN-32. J. Biol. Chem. 268 (1993) 349-355. [PMID: 8416942]
2. Oda, K., Ogasawara, S., Oyama, H. and Dunn, B.M. Subsite preferences of pepstatin-insensitive carboxyl proteinases from prokaryotes: kumamolysin, a thermostable pepstatin-insensitive carboxyl proteinase. J. Biochem. 128 (2000) 499-507. [PMID: 10965051]
3. Oyama, H., Hamada, T., Ogasawara, S., Uchida, K., Murao, S., Beyer, B.B., Dunn, B.M. and Oda, K. A CLN2-related and thermostable serine-carboxyl proteinase, kumamolysin: cloning, expression, and identification of catalytic serine residue. J. Biochem. 131 (2002) 757-765. [PMID: 11983085]
4. Comellas-Bigler, M., Fuentes-Prior, P., Maskos, K., Huber, R., Oyama, H., Uchida, K., Dunn, B.M., Oda, K. and Bode, W. The 1.4 a crystal structure of kumamolysin: a thermostable serine-carboxyl-type proteinase. Structure 10 (2002) 865-876. [PMID: 12057200]
5. Wlodawer, A., Li, M., Gustchina, A., Tsuruoka, N., Ashida, M., Minakata, H., Oyama, H., Oda, K., Nishino, T. and Nakayama, T. Crystallographic and biochemical investigations of kumamolisin-As, a serine-carboxyl peptidase with collagenase activity. J. Biol. Chem. 279 (2004) 21500-21510. [PMID: 15014068]